Ready-to-eat (RTE) food is becoming increasingly popular among restaurants and street sellers worldwide, including in Egypt. Although RTE products offer the convenience of fast food, they also represent a risk of bacterial infection as they are not heated further. S. aureus cross-contamination of meat products is a leading cause of food poisoning globally37. The current research explored the ubiquitous distribution of S. aureus in the analyzed RTE beef products. In the current study, 30.7% of RTE-tested samples were positive for S. aureus. Similar findings were reported previously in Shaanxi Province, China (34.4%)37 and Libya (32%)38. In comparison, the obtained results are higher than those documented by other foodborne illness studies on RTE beef products that were 11.8% in China39 and 23.4% in Zagazig city, Egypt40. Otherwise, this result was lower than stated in beef products at Qalubeya Governorate, Egypt (50.8%)41.
Certain restaurant staff members need to gain awareness of and adhere to hygienic protocols, particularly personal hygiene-related ones; others may partially or fully observe these protocols. The fact is that the majority of workers don’t wear aprons, gloves, or protective head covers. Rather, some employees eat, drink, and even smoke while they work. According to recent studies, luncheon samples eaten immediately without being heated had lower S. aureus levels than those (kofta, burger, and shawarma) previously39. In contrast, our results highlighted that the highest incidence of S. aureus among RTE beef products was in luncheon samples by 36.7% with a mean of 6.1 × 10 ± 0.4 × 10 CFU/g. This confirms inadequate sanitary practices at the time of luncheon preparation and processing.
Moreover, Food handlers may be responsible for contaminated meat with S. aureus due to cross-contamination from their hands42. Our results agreed well with those of other studies in Egypt41,43. Furthermore, Morshdy et al.43 declared that S. aureus was more prevalent in Egypt’s kofta (90%), luncheon (50%), burger (75%), and shawarma (80%). These variations in meat microbiological quality can be attributed to factors such as cooking method (frying, roasting, or grilling), raw ingredient quality, meat size and shape, cooking utensils (stew container, grill, oven), and seasoning (dressings, vegetables, herbs, spices)44.
The high frequency of S. aureus in beef products may be because certain employees follow hygiene procedures inside these restaurants, wiping down contact surfaces that are exposed to food, like tables, preparation boards, and utensils, using just a towel or some soap and water. On rare occasions, some continue to work while experiencing hand or respiratory problems like boils or coughing. Humans experience a wide range of diseases as a result of these conditions. Therefore, enhancing processing methods, monthly checks of workers and utensils for S. aureus, frequent disinfection of food contact surfaces, proper heating of beef products, and finally, strict personnel hygiene measures are all important to prevent the existence of S. aureus in RTE beef products42.
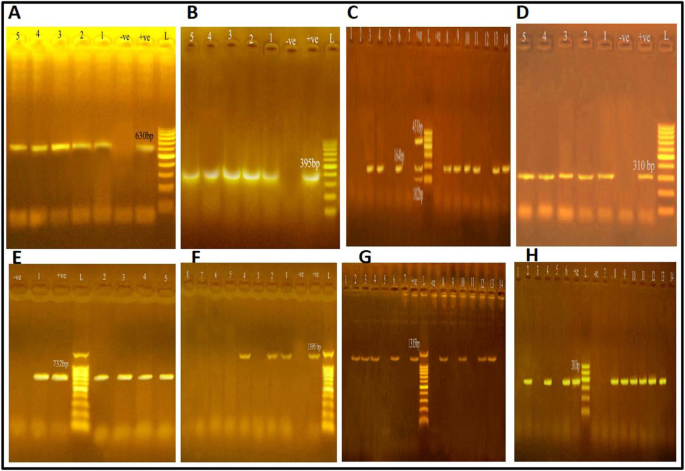
The current study declared that 58.7% of the S. aureus isolates from the samples that were looked at had coagulase, 47.6% had nuc, and 13% had only seb amongst other S.E.s encoding genes. More than 13% of S. aureus confined from RTE beef products were affirmative for more than one virulence-associated gene (Table 2). Amplification of the coagulase gene proved to be a rapid and precise approach for typing S. aureus. All explored isolates of S. aureus produce the coagulase gene, a highly virulent component. Coagulase results in plasma coagulation in the host and is an identifying marker for S. aureus infection45. The existence of coagulase and nuc genes explored in this study is comparable with previous studies 45,46.
Additionally, S.E.s are the most prominent virulence genes in S. aureus and the leading cause of staphylococcal food poisoning. The weak evidence of staphylococcal food poisoning outbreaks with the classical enterotoxins seb, sec, sed, and she can be identified commercially47. Although the Egyptian Organization for Specification and Quality Control48 requires that S.E.s not be detected in beef products, our study detected the seb gene in 13% of isolated S. aureus, which produces enterotoxins. In addition, Sallam et al. 49 identified sea, seb, and sec genes in all positive isolates. Wu et al.5 concluded that all S. aureus isolates had a minimum of one enterotoxin gene.
The incidence of S.E. genes in a wide range of isolates underlines their vertical transmission potential rather than horizontal transmission, as most of these genes exist in mobile parts of the genome. S. aureus enterotoxin explored gene differs by country due to geographical variations and is further influenced by the isolated strains’ ecological sources (food, animals, and humans). Moreover, in Poland, sec; Bulgaria, sea; and Egypt, seb are the most common toxin genes 46,50,51. The isolates’ toxin gene profiles vary according to their origin, isolation samples, and geographical location.
On the other hand, resistance to penicillin, cefoxitin, and oxacillin was significantly detected in the S. aureus isolates in this study. This finding aligns with Wu et al.5 and Abbasi et al.52, who documented an increase in β-Lactam resistance among S. aureus isolates. Also, strong resistance to aminoglycoside antibiotics was reported in previous studies, except Abbasi et al.52, who reported lower resistance to kanamycin, tetracycline, and Chloramphenicol. Moreover, Komodromos et al.53 reported similar penicillin resistance and lower Sulfamethoxazole/Trimethoprim, ampicillin, cefoxitin, tetracycline, and chloramphenicol resistance. The study found that 50% of the isolates resisted cefoxitin and were considered MRSA. This study has also revealed that shawarma had the highest occurrence of MRSA isolates, accounting for 55.5% of the samples. Following closely behind were Kofta, burger, and Sausage, each with a frequency of 50%. In contrast to the global meta-analysis, which reported a detection rate of 3.2% for MRSA in beef products, the current study reveals a significantly higher prevalence of this bacterium54. The rate of MRSA isolation observed in this study also exceeds that reported by Mahros et al.42 and Song et al. 55. However, the frequency was lower than reported by Saber et al.40.
In the current study, 15 out of 23 MRSA investigated were positive for the mecA gene, and 8 out of 23 MRSA did not show any presence of the mecA gene. These alternative mec genes, such as mecB or mecC, could be responsible for their methicillin resistance. According to Yang et al.39 and Abbasi et al.52, the prevalence of mecA was found in 6 and 32 MRSA strains, while Saber et al. 40 and Al-Amery et al. 56 amplified mecA in all MRSA strains. Furthermore, Vancomycin is the optimal therapeutic option for treating methicillin-resistant biofilm infections57. However, the World Health Organization has categorized vancomycin-resistant S. aureus (VRSA) as a “high priority” antibiotic-resistant pathogen due to its significant impact on public health. The percentage of VRSA found in our samples was higher than that found by Yang et al.39 and Al-Amery et al.56 in previously published studies. Notably, none of the isolates obtained by Wang et al.37 and Abbasi et al.52 exhibited vancomycin resistance. The varying levels of antimicrobial resistance observed across various studies may be attributed to the differences in opinions among veterinary physicians about which type of antimicrobial agents should be prescribed, the practicality of regulations that limit their use, and the pricing of such agents. These factors can significantly impact the prevalence of antimicrobial resistance within distinct geographical regions.
Eleven distinct antimicrobial resistance profiles and 39 MDR isolates were revealed. The MDR isolates observed in our study exhibit higher levels when compared to the findings reported by Velasco et al.58, and Ou et al.59. Such variations in resistance profiles can have significant implications for the treatment and control of infections caused by this bacterium. The antimicrobial susceptibility evaluation has yielded results indicating a notable occurrence of resistance towards specific antimicrobials, with a range of resistance patterns identified among the isolates of S. aureus60. This can be attributed primarily to the extensive and inconsistent utilization of these agents, particularly in veterinary medicine, resulting in a significant increase in the prevalence of antimicrobial resistance. Moreover, the excessive application of disinfectants, self-treatment with antimicrobial agents, and the provision of single-dose therapies have all contributed to this issue. Hence, it is crucial to promptly adopt rigorous preventive strategies to restrict the transmission of antimicrobial resistance across all stages of food production60.
Biofilm formation was a common characteristic among most strains of S. aureus. Our findings are consistent with previous research, highlighting the ability of S. aureus isolates from meat and meat products to form biofilms52,53. The production of biofilms by S. aureus and their simultaneous presence with saprophytic microorganisms within these biofilms undeniably gives rise to the establishment of persistent contamination reservoirs within food-processing facilities44. These findings are supported by Ou et al.59, as they revealed that food serves as an excellent adhesive medium and a reservoir for S. aureus because the common properties among food substrates, particularly their viscosity, play an overwhelmingly significant role in the successful colonization of S. aureus as compared to the differences in food surface properties and bacterial species.
The role of the ica genes in forming biofilms by S. aureus strains has been extensively explored. It has been consistently observed that the majority of biofilm-producing S. aureus strains in the current study possess the icaA and icaD genes either separately or in combination, with the icaD being predominant, which disagrees with the study of Saber et al.40 where the icaA was the most prevalent demonstrates a significant correlation with the formation of biofilms. The results obtained in this study align with Tang et al.61, who found that 87.5% of S. aureus strains isolated from different food sources possessed the icaA and icaD genes. This indicates a consistent presence of these genes in S. aureus strains across various origins. Furthermore, Abbasi et al.52 detected icaA and icaD genes in biofilm-producing S. aureus obtained from raw meat and meat products (80.4% each). These findings emphasize the importance of the icaA and icaD genes in forming biofilms by S. aureus, which can affect food safety.
In this study, isolates containing the icaD gene produced biofilms that ranged from moderate to strong. In contrast, isolates that expressed both icaA and icaD exhibited strong biofilm (p < 0.05). Mubarak and El-Zamkan62 confirmed these results. Contrarily, it has been ascertained that three isolates were identified to have solely the icaA gene and were linked with a weak biofilm phenotype. A study conducted by Gerke et al.63 explained the link between genotype and phenotype. They found that icaA alone had a weak N-acetylglucosaminyl transferase activity, but when icaD was co-transcribed with icaA, it had full activity. Biofilm is one of the useful tools used by S. aureus to resist antibiotics59, which illustrates the significant relationship between biofilm formation and MDR in this study.
The study’s findings are quite alarming as they reveal that a significant number of MRSA strains (60.9%) are also VRSA, a major cause of concern. Moreover, 21.7% of these strains have developed resistance against both vancomycin and Linezolid. These results highlight the need for urgent action to address the escalating concern of antibiotic resistance and to find new ways to combat these pathogens. Also, this study reveals that all MRSA are MDR and biofilm producers, except three isolates, non-biofilm producers, while 14 out of 18 VRSA (77.8%) are biofilm producers. S. aureus biofilm-associated infections give rise to severe and potentially fatal diseases, including endocarditis, septic arthritis, cystic fibrosis, and recurrent infections in both human and animal communities64. This finding underscores the significant impact of MRSA as an opportunistic pathogen in nosocomial infections, as it demonstrates high levels of resistance to various antibiotics and possesses a wide range of virulence factors. What adds to the concern is that a considerable proportion of MRSA strains (60.9%) also exhibit resistance to vancomycin, a major cause of concern. Moreover, 35.7% of MRSA and VRSA strains resist Linezolid, which is crucial in MRSA treatment. Another problem is that 4 MRSA/VRSA isolates harbored the seb gene. Identifying beef products as a reservoir for virulence and antibiotic-resistant S. aureus highlights the pressing need for prompt intervention. These findings emphasize the urgency of taking action to mitigate the issue of antibiotic resistance and to discover alternative methods for combating these organisms.
The 32.6% MRSA confirmation rate in ready-to-eat meat products represents a significant public health threat, particularly for vulnerable populations including hospitalized patients, immunocompromised individuals, and elderly consumers. The presence of virulent MRSA strains carrying both resistance (mecA) and virulence (nuc) genes in commonly consumed foods highlights the urgent need for integrated surveillance and control measures spanning the entire food-to-healthcare continuum9,11.
Protecting human health from the harmful consequences of hostile S. aureus has become increasingly challenging. The chemical preservatives’ destructive effects on human health, microbial resistance, toxicity, susceptibility, and their limited application have upsurged the demand for potentially effective, safer, healthy, and natural antimicrobial agents with a unique force against pathogens, especially S. aureus12. Thus, the antibacterial activity of algal extracts can provide a crucial component of S. aureus infection remedy that traditional antibacterial agents cannot. In addition, they can be used as organic preservatives to safeguard healthier and safer food away from the risks and harmful side effects of ordinary chemical preservatives12. In the current study, three algal species (crude extracts), including H. opuntia, J. rubens, and C. racemosa, exerted strong bactericidal activity, dose-dependently, against S. aureus according to seaweed species and used solvent. Persuasive antibacterial activity was recognized for C. racemosa in methanol extract against S. aureus, resulting in a reduction of serious bacterial growth (Δabs 620) at a 1.5 mg/ml concentration. However, the concentrations of 1.5 mg/ml of J. rubens (ethyl acetate extract) and H. opuntia (acetone extract) showed much less bacterial growth reduction (Δabs 620) in S. aureus (Fig. 7).
Diverse studies explored the antibacterial potential activities of marine seaweed, viz., J. rubens, H. opuntia, and C. racemosa. Different solvents, viz., petroleum ether, acetone, chloroform, methanol, ethyl acetate, ethanol, hexane, and water, were practiced for algae extraction to explore the antimicrobial algal extract activity against bacterial cultures, including S. aureus12,65. C. racemosa (methanol extract) exhibits the strongest activity against the growth of MRSA (methicillin-resistant S. aureus) with the highest antimicrobial effect (97.7 ± 0.30%)58. Similar results were obtained by Chan et al.66, using the broth microdilution technique, where methanolic, hexane, acetone, chloroform, ethyl acetate, and ethanolic extracts of C. racemosa showed different activity against Gram-positive bacteria growth (S. aureus) with a 0.36 mm inhibition zone.
Furthermore, the C. racemosa extract exhibits a dual mechanism of action against S. aureus, combining direct bactericidal effects with targeted virulence suppression. The profound inhibition of nuc and coa genes detected by RT-PCR analysis suggests the extract disrupts both immune evasion (via nuclease suppression) and clotting mechanisms (via coagulase reduction). While the log reduction demonstrates killing capacity, the virulence gene suppression occurs even at sub-lethal concentrations, potentially reducing selective pressure for resistance. This combined approach—simultaneously decreasing pathogen load and disabling virulence factors—may prove more effective than conventional antibiotics. Future studies should identify the active compounds responsible for these effects and evaluate their efficacy in animal infection models.
Moreover, it was reported that C. racemosa exhibited promising antimicrobial activity against human and food pathogens67,68. The promising efficacy of the marine seaweed C. racemosa extract against the S. aureus pathogen could be due to the active metabolites, phytochemicals, and fatty acids as well as their derivatives65,66. The promising antibacterial effects of methanol extract of C. racemosa against S. aureus could be peculiarities to the most bountiful detected phytochemicals and bioactive compounds including spathulenol (1.29%), Cubenol (0.45%), 2-Cyclohexen-1-one, 2-methyl-5-(1-methylethenyl) (0.41%), trans-calamenene (0.3%).
Additionally, in vitro data in previous literature showed that Spathulenol poses potent antimicrobial activities against diverse bacterial cultures, including S. aureus67. It has been declared that spathulenol has a major role in anti-inflammatory, immunomodulatory, and antiproliferative activities, with non-toxic, non-mutagenic, and non-tumorigenic properties, and could be a potential antimicrobial drug69. Moreover, Cubenol and 2-Cyclohexen showed antibacterial activities against S. aureus with MICs ranging from 7.81 to 15.62 mg/ml. Moreover, a promising antifungal activity against filamentous fungi was detected, which could be utilized in wide-ranging applications to prevent microbial growth as drug molecules, cosmetics, and food 70.
It is reasonable to assume that the detected compounds have potential antibacterial features and that extracts of the algae in which they are abundant have powerful antibiotic properties. The investigated algae species may be more efficient against pathogenic bacteria than typical bactericidal treatments. Hence, they may be regarded as natural preservatives, delivering nutritious and safe food through their stable, biologically active molecules without the adverse effects of chemicals. It also offers perspectives on discovering innovative antibacterial agents for food preservation or therapeutic applications.
The observed antimicrobial activity of the algal extracts can be attributed to the presence of bioactive constituents, which have been previously reported for their antibacterial properties. For instance, spathulenol has demonstrated significant antimicrobial effects71, while cubenol has been shown to inhibit bacterial growth72. Similarly, 2-cyclohexen-1-one, 2-methyl-5-(1-methylethenyl)73 and trans-calamenene74 have also been documented for their antibacterial activities. These compounds likely contribute to the overall antimicrobial efficacy of the algal extracts by targeting bacterial cell membranes, disrupting enzymatic functions, or inducing oxidative stress. The presence of such bioactive constituents in the algal extracts underscores their potential as natural antimicrobial agents.