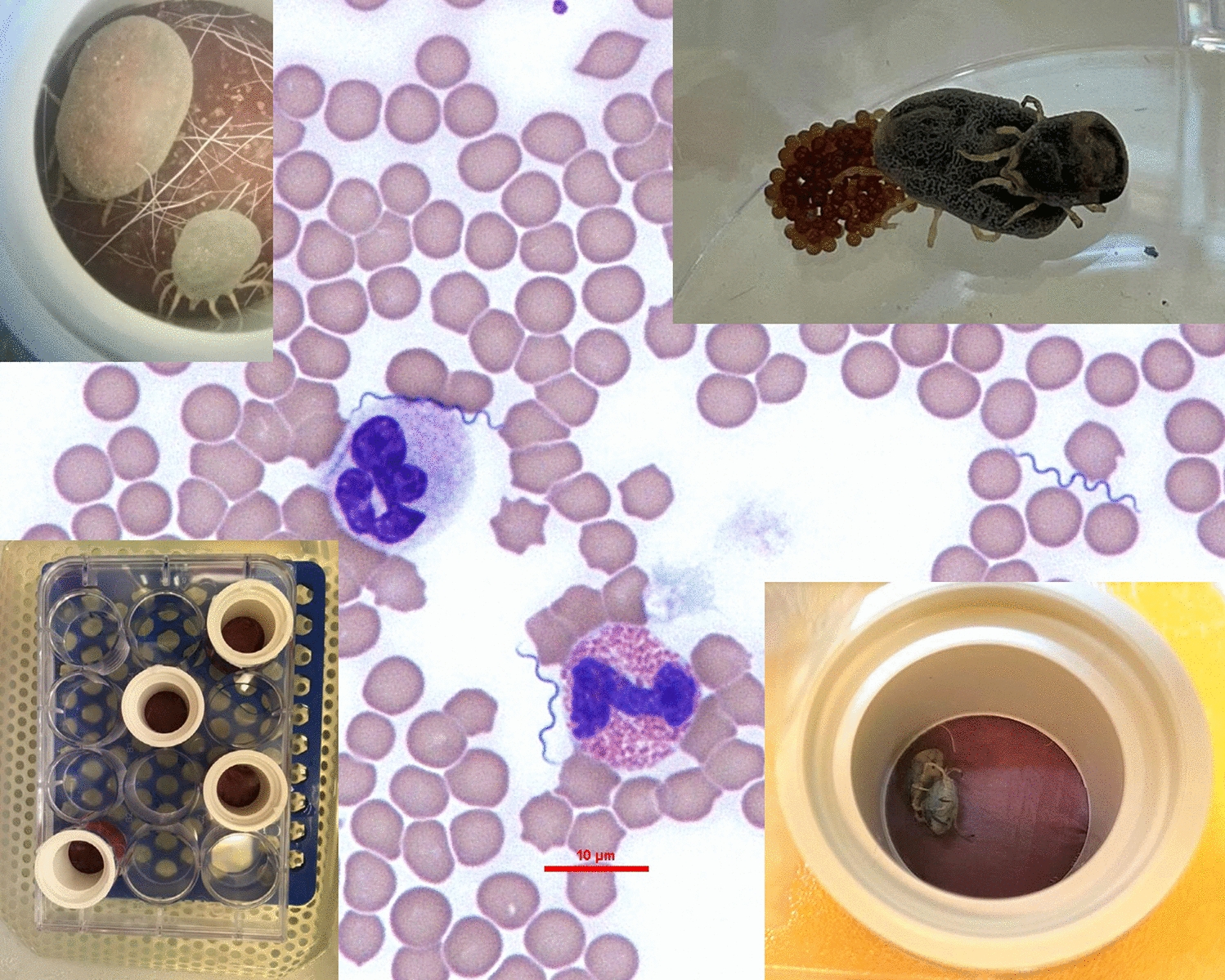
Artificial feeding and lifecycle completion
The whole lifecycle of O. tholozani ticks was completed three times in consecutive experiments using the artificial feeding system in this study. In total, 5788 ticks of different life stages were tested, including 1605 larvae, 601 first-stage, 364 second-stage, and 269 third-stage nymphs, 30 females and 34 males, and 2885 eggs. The first cycle was completed in 351 days (standard error, SE = 17.35), the second cycle in 546 days (SE = 13.8), and the third cycle in 238 days (SE = 6.15) (Additional file 1: Additional Table 1). The difference between the lengths of the cycles was due to a technical reason related to the feeding system. In the second cycle, we changed the dog from which we produced the lipid hair extract since the dog used in the first cycle was no longer available. The time between the end of one lifecycle and the next, i.e., from the first oviposition of a certain tick generation until the feeding of the first batch of larvae of the next generation, ranged between 45 and 63 days [average (AV): 51.5, SE = 5.7].
Reproductive capacity parameters, engorgement, and molting rates were measured for the different tick generations (Additional file 1, Table 1). The average number of eggs laid per female was 56.8, 183.7, 184.2, and 189.0, for the parent stock, first, second, and third generation, respectively. The percentage of larvae hatching was 60.2%, 91%, and 94% for the first, second, and third generations, respectively. The period of days between the female tick engorgement and oviposition was 44.2, 21.2, 28.7, and 26.4 for the parent stock, first, second, and third generation, respectively. Average percentage rates of engorgement were 81.9% (SE = 5.1), 58.0% (SE = 10.3), and 68.2% (SE = 2.3) for the first, second, and third generation, respectively; average molting rates were 56.9% (SE = 11), 85.6% (SE = 3.5), and 85.5% (SE = 3.9), for the first, second, and third generation, respectively. The average mortality rates of ticks in all three lifecycles were 7.1% (SE = 3.1), 4.5% (SE = 2.3), 2.4% (SE = 2.4), 12.2% (SE = 3.9), and 0%, for larvae, first-, second-, and third-stage nymph, and adults, respectively. Females were fed only once in their adult life prior to oviposition. Females, except for those of the parent stock and of the third generation, which were analyzed after the first oviposition, were allowed to continue with additional cycles of folliculogenesis for up to 5 months. Out of nine females belonging to the first and second generations, two females had four ovipositions, two had three ovipositions, three had two ovipositions, and two had only one oviposition. The other tick life stages were also fed only once in each life stage prior to molting, except for an additional feeding attempt that was carried out for those ticks that refused to feed in the first try, which was done a week after the first one. This part of the study showed that the whole lifecycle of O. tholozani can be reproduced under laboratory conditions exclusively by artificial feeding with no need for feeding on experimental animal hosts.
Borrelia persica survival in bovine blood
Follow-up of spirochete motility in the bovine blood was continued over 8 days. Motile spirochetes were observed until 96 h post-inoculation and not thereafter, supporting the idea that B. persica requires being in a tick or vertebrate host to continue its long-term survival.
Borrelia persica acquisition by third-stage nymphs at defined time points post-infection
Ticks raised in the laboratory, which originated from the parent stock, were brought from the egg stage to the third-stage nymphs by artificial feeding on uninfected bovine blood. At that stage, 42 third-stage nymphs were fed with heparinized bovine blood containing 10⁶ B. persica spirochetes per ml, and were divided into five groups according to the time point of analysis post-infection. The groups included ticks examined by PCR for B. persica in their guts and salivary glands, 1, 2, 3, 4, and 5 weeks after infection (Table 1). All 16 (100%) of the third-stage nymphs tested at 1 and 2 weeks after infection were found to be positive for B. persica by PCR only in their guts. At week 3, seven out of eight (87.5%) ticks tested were positive for B. persica in their guts, and one was negative in both guts and salivary glands. In the fourth week, two out of eight third-stage nymphs had molted to females and one to a male, while the remaining five stayed as third-stage nymphs. Six (75%) of these ticks were infected, including five third-stage nymphs and one female. When relating to all tick stages, B. persica DNA was found in the guts of four ticks and salivary glands from three. Two third-stage nymphs were positive in both their guts and salivary glands, two were positive only in their guts, and one was positive only in its salivary gland. The positive female was infected in its guts and its salivary glands. Of the ten ticks evaluated after 5 weeks of infection, three nymphs had molted to fourth-stage nymphs, and one had molted to a male. Infection was detected in five ticks, including five guts and one salivary gland. Three third-stage nymphs were positive only in their guts, one fourth-stage nymph was infected in its gut, and another fourth-stage nymph was infected in its gut and salivary glands. The male was negative.
In total, out of the 42 infected third-stage nymphs in all the groups, 34 (81%) ticks (31 third-stage nymphs, two fourth-stage nymphs, and one female) were positive for B. persica by the flaB PCR, including 33 guts and 5 salivary glands, with infection of salivary glands detected only in ticks analyzed at week 4 and 5 after infection (Table 1). The average B. persica loads in the guts, expressed as Borrelia flaB copies per tick 16SrRNA gene copy, ranged between 4.7 and 67.4 in the different weeks (Table 1). The copy number of B. persica in the guts at week 1 (AV = 67.4) was significantly higher than at week 3 (AV = 5.5) and 4 (AV = 48.7) (Kruskal–Wallis test H = 8.25, df = 3, P = 0.041; Table 1). However, when comparing the B. persica loads in the guts of ticks at week 5 of infection (AV = 28.5) with those found at the other time points, only a nonsignificant trend was observed (Kruskal–Wallis test H = 8.79, df = 4, P = 0.066). The average B. persica loads in the salivary glands at weeks 4 and 5 post-infection were 7.9 flaB-copies per tick 16SrRNA gene copies (SE = 7.5), and in the guts analyzed at week 4 and 5 it was 37.46 (SE = 23.43) with no significant differences between loads in the salivary glands and in the guts with those found at the other time points (Mann–Whitney U-test: U(13) = 16.00, Z = −0.309, P = 0.825). The results described in this section portray the dynamics of B. persica within experimentally infected ticks, with movement of infection from the tick gut to the salivary glands noted from 4 weeks post-infection.
Transstadial and transovarial transmission of B. persica
Transstadial transmission was analyzed by infecting 161 larvae with 1.2 × 10⁷ B. persica spirochetes per ml of blood and testing 20 individuals from each tick life stage, e.g., larvae, first-, second-, and third-stage nymphs, and adult ticks (ten females and ten males). Larvae were tested 1 week after feeding on infected blood, whereas first-, second-, and third-stage nymphs were analyzed a week after molting; adults were analyzed 2 weeks after molting. Except for larvae, which were analyzed after engorgement, the remaining stages were analyzed unengorged. The rate of infection decreased from 100% (20/20, 95% CI 83.16–100%) in larvae to 55% (11/20, 95% CI 31.53–76.94%), 20% (4/20, 95% CI 5.73–43.66%), and 25% (5/20, 95% CI 8.66–49.10%) in first-, second- and third-stage nymphs, respectively. Adult ticks showed an infection rate of 20% (4/20, three male ticks and one female tick).
Average B. persica loads in ticks were 243.8 (SE = 42.1), 21.6 (SE = 7.7), 38.1 (SE = 4.9), and 61.2 (SE = 21.9) flaB-copies per tick 16SrRNA gene copies in larvae and first-, second-, and third-stage nymphs, respectively. Average B. persica loads in adults were 199.2 flaB-copies per tick 16SrRNA gene (SE = 25.7) [244.4 (SE = 50.2) and 63.5 in males and females, respectively] (Table 2). There was a significant difference in parasite load between tick stages (Kruskal–Wallis test H = 20.18, df = 4, P < 0.001). Post hoc pairwise comparisons using Dunn’s test with Bonferroni correction indicated that the larvae had a significantly higher parasite load than first-stage nymphs (Z = 18.344, P = 0.002), but no other pairwise differences were significant.
Transovarial transmission was tested by mating eight infected females with eight uninfected males and analyzing 20 eggs and 20 larvae laid and hatched from each of the eight potentially infected females. Because only eight eggs were analyzed from one of the females, a total of 148 eggs and 160 larvae were analyzed. The number of days from feeding and engorgement of the females to the beginning of the first oviposition ranged between 21 and 44 days (average = 26.6, SE = 2.6), and the number of eggs laid per female ranged between 70 and 190 (average = 129.6, SE = 13). Larval hatching rate ranged between 67% and 85% (average = 77%, SE = 2.3). Dissection and molecular analysis of the females and males, which were infected at third-stage nymph, was done after the first oviposition between 12 and 16 weeks post-infection, and real-time PCR targeting the Borrelia flaB gene [21] was done on the salivary glands, guts, and gonads. Of the eight females, five were infected (62.5%). Three of them were positive in their guts, five in their salivary glands, and three in their ovaries. Of those, three females were positive in their guts, salivary glands, and ovaries, and two females only in their salivary glands. All eight uninfected males that were mated with the females were negative for B. persica DNA. Regarding B. persica infection in eggs, 4 of 148 eggs (2.7%) analyzed were positive by PCR. The positive eggs belonged to three positive females. Considering only the eggs laid by infected females (n = 5), the rate of egg infection was 4 of 100 (4%), ranging from 5% to 10% per infected female (Table 3). Regarding B. persica infection in larvae that hatched from the eggs of the experimentally infected ticks, 2 of 160 larvae tested were positive (1.3%), and both belonged to larvae hatched from eggs laid by one positive female. Considering only larvae derived from positive females, the rate of larvae infection was 2 of 100 (2%) and 10% per the single positive female, which gave rise to the infected larvae (Table 3).
Five uninfected females were paired with five uninfected males as controls. The period of days from feeding and engorgement of the adults until the beginning of the first oviposition ranged between 21 and 29 days (AV = 26.4, SE = 1.6), and the number of eggs laid per female ranged between 84 and 263 (AV = 189, SE = 29.3). The larvae hatching rate ranged between 74% and 94% (AV = 88%, SE = 3.9%). The hatching rate of the larvae originating from the control group of uninfected females (88%) was close to being significantly higher than the hatching rate of the larvae derived from B. persica-infected females (75%) (Welch’s t-test, t(5.477) = −2.49, P = 0.051).
The outcomes of the experiments on transstadial and transovarial transmission indicated that both modes of B. persica transmission could be demonstrated by experimental infection using artificial tick feeding, and that the rate of transovarial transmission was considerably lower than that of its transstadial counterpart.
Horizontal transmission of B. persica between ticks by hyperparasitism
Twenty-two third-stage nymphs infected with heparinized bovine blood containing 10⁶ spirochetes per ml were divided into two groups. In the first group, each infected third-stage nymph was individually paired immediately after infection with an uninfected unfed male for 3 h to study possible direct acquisition of B. persica by the males from the nymphs. Biting of nymphs and engorgement of the males with laceration of nymph dorsal plates caused by male mouthparts were observed in three nymph-male pairs (Figs. 2A, B). Five weeks after pairing, three out of ten potentially infected and engorged nymphs molted into fourth-stage nymphs, one molted into a male, and the rest remained as third-stage nymphs. Molecular screening by real-time PCR targeting the flaB gene [21] revealed that five out of these ten ticks were positive for B. persica, including three third-stage and one fourth-stage nymph. All positive ticks showed infection in the guts, and one fourth-stage nymph showed B. persica DNA also in the salivary glands. Out of ten uninfected, unfed males before the pairing, three showed blood in their gut during their dissection, which was performed 5 weeks after pairing with the nymphs. These three males had been paired with third-stage nymphs, which showed laceration lesions in their dorsal plates after the pairing, and two of these males were positive for B. persica flaB DNA in their guts at week 5 of the experiment. These two positive males had been paired 5 weeks earlier with two nymphs that were PCR-positive, demonstrating transmission of B. persica by hyperparasitism. The third engorged male, which was negative by PCR, was paired with a nymph that turned out to be PCR-negative for B. persica when tested.
Laceration lesion in a nymphal dorsal plate (black arrow) immediately after hyperparasitism by a male (A), and healed laceration lesion in the dorsal plate (black arrow) of the same nymph 5 weeks after the breach caused by a male (B)
In the second group, 12 uninfected, engorged third-stage nymphs were paired individually immediately after feeding with 12 potentially infected unfed males for 3 h. After the pairing, five nymphs showed laceration of the dorsal plates and one on the ventral exoskeleton, caused by male mouthparts. Five weeks after pairing, 3 out of 12 engorged uninfected third-stage nymphs molted into males, 1 into a female and 1 into a fourth-stage nymph, and the remaining 7 remained as third-stage nymphs. Borrelia-specific flaB DNA real-time PCR screening revealed that all of the previously engorged uninfected ticks were negative for B. persica DNA, while two of the potentially infected males, which fed on the uninfected ticks, harbored B. persica DNA in their guts. In essence, B. persica was not transmitted during hyperparasitism from infected males to engorged nymphs. The results of this part of the study indicate that B. persica can be transmitted between O. tholozani ticks by feeding directly on each other.
Molecular analysis
All positive samples were sequenced. In total, 101 flaB DNA sequences of B. persica were obtained. From these, 38 sequences were from 38 ticks infected during the transstadial infection experiment, 38 sequences were from positive organs of 34 ticks infected weekly during 5 weeks, 17 sequences were from positive organs and offspring of infected females during the transovarial infection experiment, and 8 sequences were from organs of infected nymphs and males that participated in the hyperparasitism infection experiment. All DNA sequences were 100% identical to each other. Four DNA sequences were submitted to GenBank (accession nos. MW284983-86). Uninfected control ticks of all life stages and all DNA extractions from negative controls were PCR-negative for B. persica.