As a lonely, bedridden child, Ory Yoshifuji wanted nothing more than a second body he could use to attend school and see his friends.
For three and a half years, sickness forced Ory to stay at home, his bedroom becoming his prison.
“All I could do was stare at the ceiling day after day. The stress of loneliness at that time became so unbearable,” he says.
“I wondered, why couldn’t I have a second body?
“I might have been able to attend school with the other body.”
Years later — after studying robotics at university — he turned his dream into a reality by opening Dawn Avatar Robot Cafe in Tokyo, where cute robot avatars welcome and entertain guests, as well as serve drinks and food.
Ory Yoshifuji fulfilled his childhood room of creating a second body for himself through robot technology. (ABC News: James Oaten)
But unlike the competition, these robots are not powered through artificial intelligence.
Instead, they are operated by real people, housebound and disabled, just like Ory was.
“We’ve managed to create a situation where people, even if they can’t physically move around much, can still participate as members of society and as productive members of the workforce,” Ory says.
‘A role in society’
The robot café has been a hit, growing from about 10 to 100 robot operators, known as pilots.
Each pilot moves the robot through a specialised program on their home computer, and can see and speak to guests.
Customers are served by people-operated robots at Dawn Avatar Robot Cafe. (ABC News: James Oaten)
The project has now begun a new venture, tapping into Tokyo’s tourism boom.
Instead of just serving guests at the café, customers can also carry a robot on their back for a personalised tour of the local area.
Ory says the idea came after many of the robot pilots joked about escaping the café.
“[The robots] approach the automatic doors, head outside, and the wi-fi signal would cut out,” he says.
“They’d often play around like that as a joke.
“But, of course, they actually wanted to go outside too.”
One pilot is Machun, who has multiple sclerosis.
Machun started working at the cafe after losing her previous job at a bank. (ABC News: James Oaten )
She lost her job at a banking think tank when her condition worsened after contracting COVID.
“I couldn’t stay awake,” she says.
“I used to be able to walk, but [then] I couldn’t do that either.”
For her, working as a robot pilot has given her a second life. Her mental health has flourished, she adds.
“Someone who could barely sit for an hour can now work six or seven hours a day with breaks,” she says.
“I now feel like I have a role in society. I really feel that.”
It is not just the pilots who enjoy the new-found freedom, but the tourists too.
Canadian tourists in Tokyo. (ABC News: James Oaten )
“What I loved about having our tour guide right on our shoulder is I feel like it was really flexible,” Canadian tourist Andrea Wheaton says.
“Especially with having young kids.”
Her husband, Dave Schultz, enjoyed the human aspect.
“You can hear the warmth through their voice and some giggles on their end,” he says.
‘Working while teleporting’
The rise of artificial intelligence and robotics has sparked debate about how such technology may destroy jobs and lead to fewer connections between people.
Ory is proud that his technology is doing the opposite.
“We are creating a society where we can live without needing to ask anyone for help,” he says.
“This also means we are creating a society where no-one is needed.”
He hopes to expand the robot tour guides to cover more areas of Tokyo.
But one of the biggest challenges is that crowded hotspots can have patchy internet, which would disrupt the robot’s connection to the pilot.
A worker operates a robot that works at Dawn Avatar Robot Cafe in Tokyo. (ABC News: James Oaten )
“I hope we can create this way of working in other locations, or even beyond Japan,” Orly says.
“It enables working while teleporting.”
And that, he adds, would help create a “society without loneliness”.
HIP HIP! HUZZAH! We are excited to host our 4th annual Renaissance Faire Night on Sunday, January 18! Before the game from 3-5pm, join us on the East Plaza for Renaissance Faire activities such as duels…
Several of Carnival Corporation’s eight world-class cruise lines made a big splash at this year’s Travel Weekly Readers Choice Awards, earning top honors across a variety of categories that celebrate excellence in cruising. From short itineraries to world voyages, from tech innovation to advisor programs, trusted travel advisors who are instrumental to the industry’s success chose winners based on the cruise lines’ excellence in service, product, and overall performance.
We proudly took home industry-best recognition in the following categories:
Best in Cruise Lines
Domestic: Carnival Cruise Line
Alaska: Princess Cruises
Under 1,000 Berths: Seabourn
World Cruise Itinerary: Holland America Line
Group Program: Carnival Cruise Line
Shipboard Tech: Princess Cruises
Short Itinerary: Carnival Cruise Line
Transatlantic Sailing: Cunard
Best Travel Advisor Educational Program
Best Advisor Loyalty Program
These awards are a testament to the incredible experiences our cruise lines deliver every day – and to the dedication of our teams and partners who make it all possible.
For beer brewer Richard Jeffares, his business is personal.
After a diagnosis of coeliac disease, the craft beer enthusiast mourned that he could no longer have a schooner at the pub with friends, given that a key ingredient in most traditional beers, barley, contains gluten.
“Somebody said to me, ‘Why don’t you start a gluten-free brewery?’” Mr Jeffares recalled.
Eight years on, the businessman is on a mission to “ensure that every liquor licence in the country offers their consumer a gluten-free beer”.
To grow its sales team to achieve that dream, his Victorian company raised about $2.5 million from 993 investors in a matter of days through a method called crowd-sourced funding, also known as equity crowdfunding.
As the name suggests, it works like a crowdfunding platform (think Kickstarter or GoFundMe), but instead of a donation, contributors invest.
It’s a way for smaller, private companies that aren’t listed or floating on the stock exchange to raise money by selling shares during a limited campaign period.
CSF, as it’s known, has only existed in Australia since 2017.
In the age of social media, it’s a method that thrives on brand strength and popularity.
Many of those who tipped into Mr Jeffares’s company, TwoBays Brewing, are diehard fans and keen drinkers of the beers themselves.
To Mr Jeffares, they’re not just investors but “brand ambassadors for us all around the country”.
“I had a gentleman from Orange yesterday giving me leads into two venues in Orange and in New South Wales,” he said.
“So they are [trying to] get our product – and I, when I talk to them, I tell them, ‘It’s your product now as well’ – into local venues so they can have a beer with their friends.”
A bid to bust business archetypes
Kirstin Hunter, the CEO of Birchal – one of the larger CSF platforms in Australia, the other being OnMarket, which carried the TwoBays campaign – champions crowd-sourced funding as an equaliser in the stereotypically male-dominated world of start-up businesses and the firms that fund them.
Australia, she says, has “a bit of a problem in the venture capital ecosystem when it comes to allocating funding to founders who are outside of a particular kind of narrow archetype”.
Birchal chief executive Kirstin Hunter. (ABC News: John Gunn)
As an example of the difficulties some entrepreneurs face, Birchal detailed a recent campaign for sex toy brand Normal Co.
Its founder, Lucy Wark, says the business couldn’t hurdle investment house internal policies “designed to restrict socially harmful investments” even though the company aimed to have a positive social impact.
“The free online sex education resources which are funded by our toys have now been viewed tens of millions of times in over 40 countries,” Ms Wark said.
“But when we actually passed the investment committee at a major VC fund, we ended up being blocked by vice clause concerns.”
According to Birchal’s annual report on the Australian crowdfunding landscape, 32 per cent of all crowdfunding capital went to teams with at least one woman founder, more than double the 15 per cent representation seen in venture capital.
The report also shows that the food and beverage sector is the largest in terms of funds raised this way.
Richard Jeffares’s company TwoBays is one of many alcohol brands, including craft brewery Philter and distillery Prohibition Liquor, to bring their fans on board.
But Birchal reports the sector has been volatile since the 2022 tech boom, when total funds raised across all platforms peaked at $86 million.
It suffered a crash in the 2024-25 financial year, falling from $65 million in the previous period to $33 million.
Kirstin Hunter said that echoed a tightening across the broader venture capital landscape due to the cost of living, but that the second half of the 2026 financial year was “looking much, much stronger”.
Meanwhile, not every crowdfunding campaign ends with a success story.
Big raises don’t always translate to big business
When Clinton Schultz launched a crowdfunding campaign for his alcohol-free brewery Sobah, he said he had the best of intentions.
As an Indigenous business owner seeking backing for his brand, which utilises native ingredients, he said it was difficult to “attract mainstream investment (and] get non-Indigenous entities to believe in and back” his business.
“Equity crowdfunding gives people an opportunity to dip their toes in (to investing) in a way that’s reasonable and affordable,” he said.
Sobah co-founder Clinton Schultz. (Supplied )
Sobah’s campaign raised more than a million dollars from the brand’s supporters.
But Mr Schultz said the notoriously competitive craft beer landscape – “dominated by the duopoly” of beer giants Kirin and Asahi – brought Sobah undone, with the company going into administration in September.
“It’s been very stressful,” Mr Schultz said.
“We’re trying to find a way forward.“
He said he was hopeful of reviving the company.
But in the meantime, it’s not yet clear how much creditors will get back, including those who tipped into Sobah’s crowdfunding campaign.
This year, Australian Distilling Co. also went under and has been forced to put its South Australian property – where it had plans to build a “Cathedral of Gin” under its Old Young’s brand – up for sale.
The company had crowdfunded nearly $2.7 million to support the project, but administrators anticipate that those unsecured investors are likely to receive “approximately 3 to 17 cents in the dollar”.
It comes after the demise of Zero Co, the laundry and soap company which closed this year after six years in business and Australia’s largest crowd-sourced raise to date, which hauled in $5 million.
“Any kind of investing is always risky, and crowd-sourced funding is no exception,” Kirstin Hunter said.
“As the licensed intermediary, we remind them on multiple occasions, and the crowdsourced funding offer document has a long risk disclosure as well.”
Buyer – and business – beware
Daniel Eason is the accounting director at BlueRock, a firm that previously worked for Birchal and continues to help businesses run crowd-sourced funding campaigns.
He believes CSF can work well for businesses, provided companies have made the right preparations.
BlueRock accounting director Daniel Eason. (ABC News: Patrick Stone)
That includes locking in 60 to 70 per cent of their fundraising target before the campaign even goes live – a task that involves some serious networking.
Then there are greater accounting costs to manage.
“The minute you go CSF, you effectively become an unlisted public company (in that you have] obligations under the corporations act to prepare general purpose financial statements and have them audited,” he said.
“Everyone thinks the raise is going to blow their socks off … but they’re [also] putting themselves into a higher cost environment going forward.”
Mr Eason said it was “the price of success, with his advice to companies looking to raise funds using CSF: go big or go home.
“The real shame is the ones who go into it who only raise 200k and go insolvent two years after,”
he said.
He also said there was a level of “buyer beware” for people looking to invest in crowdfunding campaigns.
Unlike buying shares in the more traditional way on the stock market, it often takes longer for CSF investors to see a dividend after they invest, as the companies tend to reinvest profits.
“They deploy the capital to buy that new bottling line … until they can [eventually] build that market share, make a profit and start paying dividends,” Mr Eason explained.
It can be appealing for people looking for a quick and easy way to invest, he said, “but the liquidity is also not there if, in two years, they need to sell those shares.
“That’s why CSF works better for brands that you know and love, or products that you can get behind, or industries that you understand.”
TwoBays Brewing founder Richard Jeffares. (ABC News: Darryl Torpy)
Richard Jeffares said his crowdfunding campaign took years of careful calculations, waiting for the business to reach the right level of growth, securing support in advance of the raise, and heavy marketing investment.
“It’s not a cheap exercise to go through this, from dealing with PR companies, dealing with the intermediary, and campaign managers,” he said.
“So we really felt we wanted to lean into it strongly and ensure that we didn’t leave any stone unturned.”
He acknowledges that achieving his dream of a gluten-free beer in every Australian watering hole could mean one day being bought by a larger company – which would see shareholders take their cut – but he believes his business has more room to grow first.
“We hope that these investors coming on board are proud of their shareholding and that with that growth in the business, we’re able to give them a return at some point in the future.”
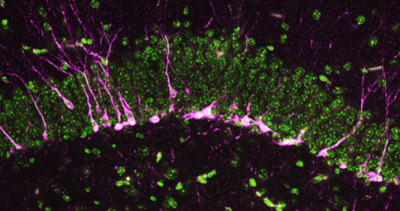
Newswise — Scientists at Johns Hopkins Medicine say results of a new National Institutes of Health-funded study are advancing efforts to exploit a new target for Alzheimer’s disease: a protein that manufactures an…
You don’t have permission to access “http://www.mountsinai.org/about/newsroom/2025/mount-sinai-health-system-ceo-named-to-modern-healthcares-100-most-influential-people-in-healthcare” on this server.
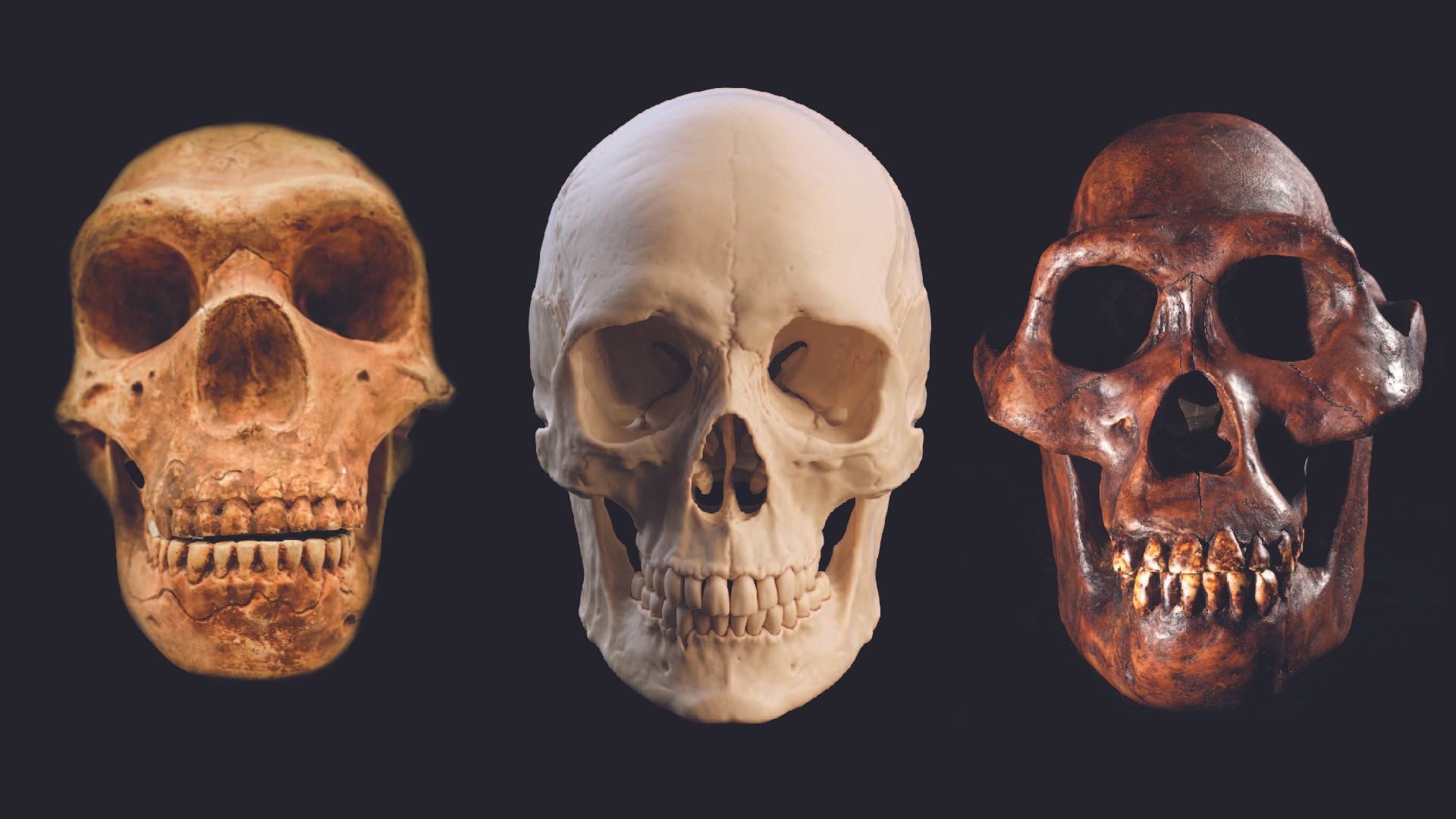
Neanderthals have fascinated scientists since they were first discovered in the 19th century. Their long heads and low brow ridges initially convinced experts that Neanderthals were some kind of evolutionary wrong turn that ended up in European…