Researchers at Tokyo Metropolitan University have uncovered the real reason liquid drains from foams, resolving a long-standing scientific puzzle. Traditional physics models have consistently overestimated how tall a foam must be before liquid…
Author: admin
-

DNA reveals two new bass species hidden in plain sight
University of Georgia researchers have identified and formally described two previously unrecognized species of black bass, known as Bartram’s bass and Altamaha bass, in a recently published study.
Although the species are only now being…
Continue Reading
-

A surprising foam discovery could change everyday products
Researchers at Tokyo Metropolitan University have uncovered the real reason liquid drains from foams, resolving a long-standing scientific puzzle. Traditional physics models have consistently overestimated how tall a foam must be before liquid…
Continue Reading
-

Metaverse inventor Neal Stephenson says VR goggles are dead • The Register
Science fiction author Neal Stephenson, who coined the term “metaverse” in his 1992 novel Snow Crash, has argued he and others who believed immersive environments would require head-mounted hardware got it wrong.
In a post penned to mark…
Continue Reading
-

Gold price in Pakistan for today, March 24, 2026 – Profit by Pakistan Today
- Gold price in Pakistan for today, March 24, 2026 Profit by Pakistan Today
- Gold falls over Rs43,000 to Rs447,762 per tola Dawn
- Gold price per tola plunges record Rs43,600 in Pakistan Business Recorder
- Gold plunges below Rs450k The Express Tribune
- Gold Rate Drops by Nearly Rs. 50,000 in Pakistan ProPakistani
Continue Reading
-

‘Stop this savage being’: Iranians fear postponed Trump attack is merely disaster delayed | US-Israel war on Iran
A wave of temporary relief, and some jubilation swept through Iran as Donald Trump announced he was postponing an attack on its energy infrastructure after he claimed to have had productive conversations with Tehran – conversations Iran…
Continue Reading
-

Capella at Galaxy Macau will change everything you know (for the better) about Asia’s most misunderstood destination
Opting for quality over quantity, and boasting some of the most mind-blowingly beautiful hotel suites we’ve ever seen, Capella at Galaxy Macau
For the better part of the last two decades, Macau has defined itself by scale and excess:…
Continue Reading
-
Pakistan Day commemorated at Pakistan High Commission, Ottawa – RADIO PAKISTAN
- Pakistan Day commemorated at Pakistan High Commission, Ottawa RADIO PAKISTAN
- AJK marks Pakistan Day with rallies, renewed pledge for Kashmir freedom Dawn
- The dream that became a nation Geo News
- Pakistani community marks National Day with fervour…
Continue Reading
-
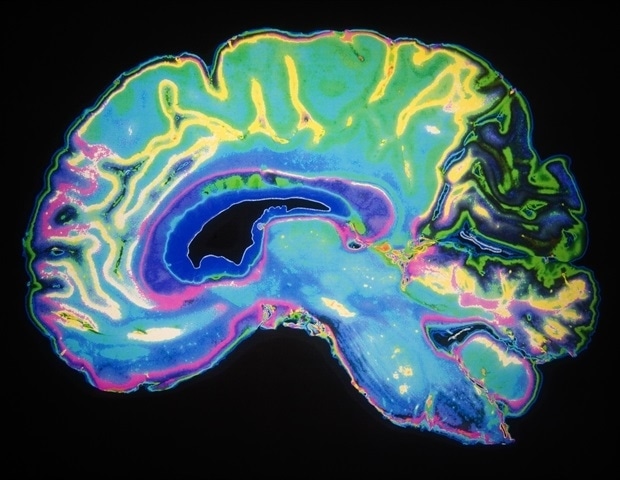
Scientists discover similarities in brain aging between mice and humans
By scanning the brains of mice throughout their lifespans, scientists at Columbia’s Zuckerman Institute and the University of Texas at Dallas have discovered that the human brain is not unique in how it changes with age. These…
Continue Reading
-
Study revises the understanding of neutrophil dynamics during respiratory viral infection
Neutrophil infiltration is a hallmark of pulmonary inflammation during respiratory viral infections, yet the origin of these cells has been a subject of debate. Using a golden hamster model of SARS-CoV-2 infection, the study led by…
Continue Reading