Here’s something exciting for everyone who loves looking at the sky! On March 3, 2026, the Moon will move completely into Earth’s shadow, making a total lunar eclipse. During this, the Moon will look deep red, which is why people call it a…
Category: 7. Science
-

Prosthetic arm blurs the line between machine and human body
A virtual forearm can bend in a blink. It can also take its time, easing toward a target as if it is thinking about the move.
In a new virtual reality study, both extremes felt wrong.
When a prosthetic arm moves on its own, speed turns out to be…
Continue Reading
-

NASA Spotted This Comet as It Exploded, and It’s Full of Organic Chemicals
NASA’s SPHEREx space telescope has observed interstellar comet 3I/ATLAS flaring dramatically months after its closest pass by the Sun. Only the third confirmed interstellar object ever detected in our solar system, 3I/ATLAS is offering…
Continue Reading
-

Snakes keep evolving into cannibals — here’s what scientists think is going on
Cannibalism might seem like a rare and unnatural occurrence, but the behavior has arisen in several snake lineages throughout evolutionary history, often triggered by environmental stressors, scientists theorize.
When researchers reviewed 500…
Continue Reading
-
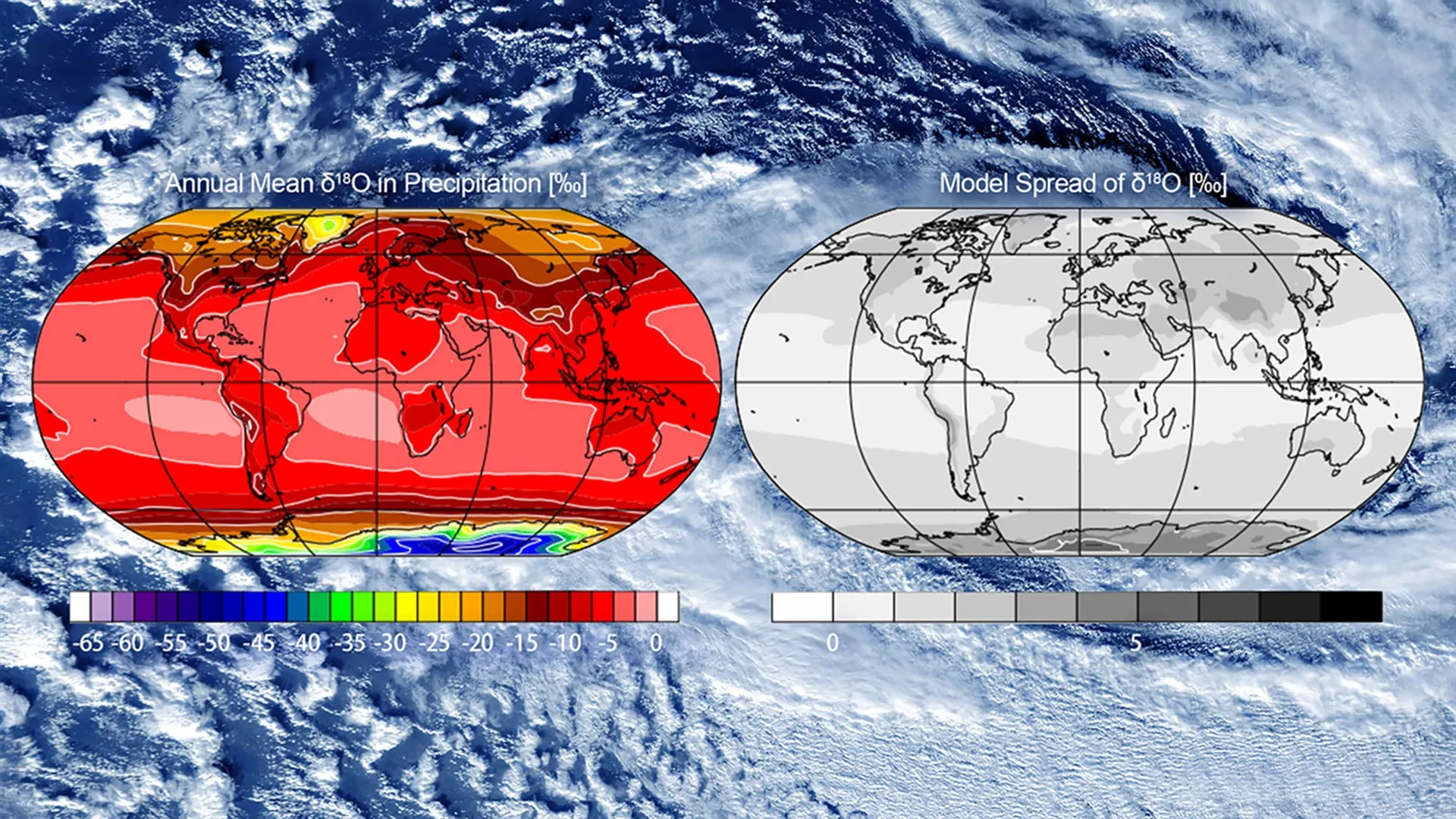
Tracking global water circulation using atomic fingerprints
Scientists can now follow the path of a single drop of water across the globe. Water is made of hydrogen and oxygen, and some of these atoms naturally exist in slightly heavier forms called isotopes. As water evaporates, forms clouds, and moves…
Continue Reading
-
Tracking global water circulation using atomic fingerprints
Scientists can now follow the path of a single drop of water across the globe. Water is made of hydrogen and oxygen, and some of these atoms naturally exist in slightly heavier forms called isotopes. As water evaporates, forms clouds, and moves…
Continue Reading
-
Ancient Rocks Reveal Hidden Climate Motion During Snowball Earth – SciTechDaily
- Ancient Rocks Reveal Hidden Climate Motion During Snowball Earth SciTechDaily
- Earth had seasons during Ice Age, according to new study BBC
- Earth’s coldest ocean conditions quantified China Daily
- Snowball Earth wasn’t fully frozen: ice-free…
Continue Reading
-

Bright green comet C/2024 E1 nears closest approach before leaving solar system
The bright green comet C/2024 E1 (Wierzchoś) displays increasing…
Continue Reading
-

Lucky few to see ‘ring of fire’ solar eclipse over Antarctica on Feb. 17
While the new moon this Tuesday (Feb. 17) will pass without much fanfare in most of the world, something more exciting will be taking place over a sliver of Antarctica: a “ring of fire” solar eclipse, also known as an annular solar…
Continue Reading
-

Long-term correlation found between Earth’s magnetic field strength and atmospheric oxygen
An analysis of geological proxy records shows that Earth’s atmospheric oxygen levels and geomagnetic field strength evolved in parallel over the past 540 million years. The study compares reconstructions of atmospheric oxygen concentration…
Continue Reading