- Scientists Just Found a Hidden Oxygen Drain Inside Plant Cells SciTechDaily
- Scientists Discover a Version of “Tug of War” Inside Cells وكالة صدى نيوز
- Scientists discover oxygen tug of war inside plant cells ScienceDaily
- Plant…
Category: 7. Science
-
Scientists Just Found a Hidden Oxygen Drain Inside Plant Cells – SciTechDaily
-

Deep sea microbial life signatures found in a very unexpected place
Far below where sunlight can reach, subtle marks in Moroccan stone suggest that life left a record in a place long assumed to be erased by time.
The finding matters because it challenges where scientists think fragile traces of early ecosystems…
Continue Reading
-

Spider uses its legs in a way never seen in any known material
Researchers have found that an Australian net-casting spider builds a hunting web that stretches far beyond ordinary spider silk and then snaps back without tearing.
That unusual elasticity allows the spider to launch a handheld net at prey and…
Continue Reading
-

Giant python snake species over 13 feet long discovered in Taiwan
Scientists have identified the first fossil evidence that a giant python more than 13 feet long once lived on Taiwan.
The discovery reveals that the island once supported far larger predators than anything found there today.
Inside one bone
Continue Reading
Bifurcation analysis and exploration of new optical soliton solutions in parabolic law medium with weak non-local nonlinearity
Zhang, Z., Yang, Z., Wang, C. & Xu, W. Accelerating ultrashort pulse laser micromachining process comprehensive optimization using a machine learning cycle design strategy integrated with a physical model. J. Intell. Manuf. 35(1), 449–465…
Continue Reading

How often do people really fart? Scientists built smart underwear to find out
Researchers at the University of Maryland have developed Smart Underwear, the first wearable device designed specifically to measure human flatulence. The small sensor tracks hydrogen in flatus, allowing scientists to reexamine long held…
Continue Reading

Curiosity Cracked Open a Rock on Mars And Revealed a Huge Surprise : ScienceAlert
A rock on Mars spilled a surprising yellow treasure after Curiosity accidentally cracked through its unremarkable exterior.
When the rover rolled its 899-kilogram (1,982-pound) body over the fragile lump of mineral in May 2024, the deposit…
Continue Reading
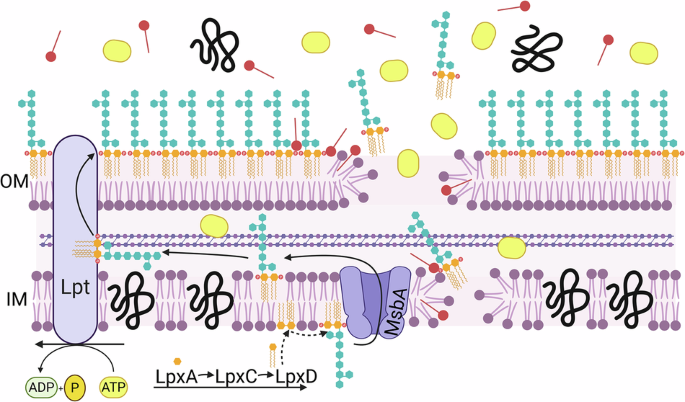
Combinations of colistin and bacteriocins prevent the selection of colistin resistance in Acinetobacter baumannii
Gigante, V. et al. Multi-year analysis of the global preclinical antibacterial pipeline: trends and gaps. Antimicrob. Agents Chemother. 68, e0053524 (2024).
GBD 2021 Antimicrobial Resistance…
Continue Reading

NASA’s DART Mission Also Changed Didymos’ Orbit Around Sun
NASA’s Double Asteroid Redirection Test (DART) spacecraft impacted the asteroid moonlet Dimorphos, which orbits the larger asteroid Didymos, in September 2022. The purpose of this mission was to test the kinetic impactor method, a…
Continue Reading
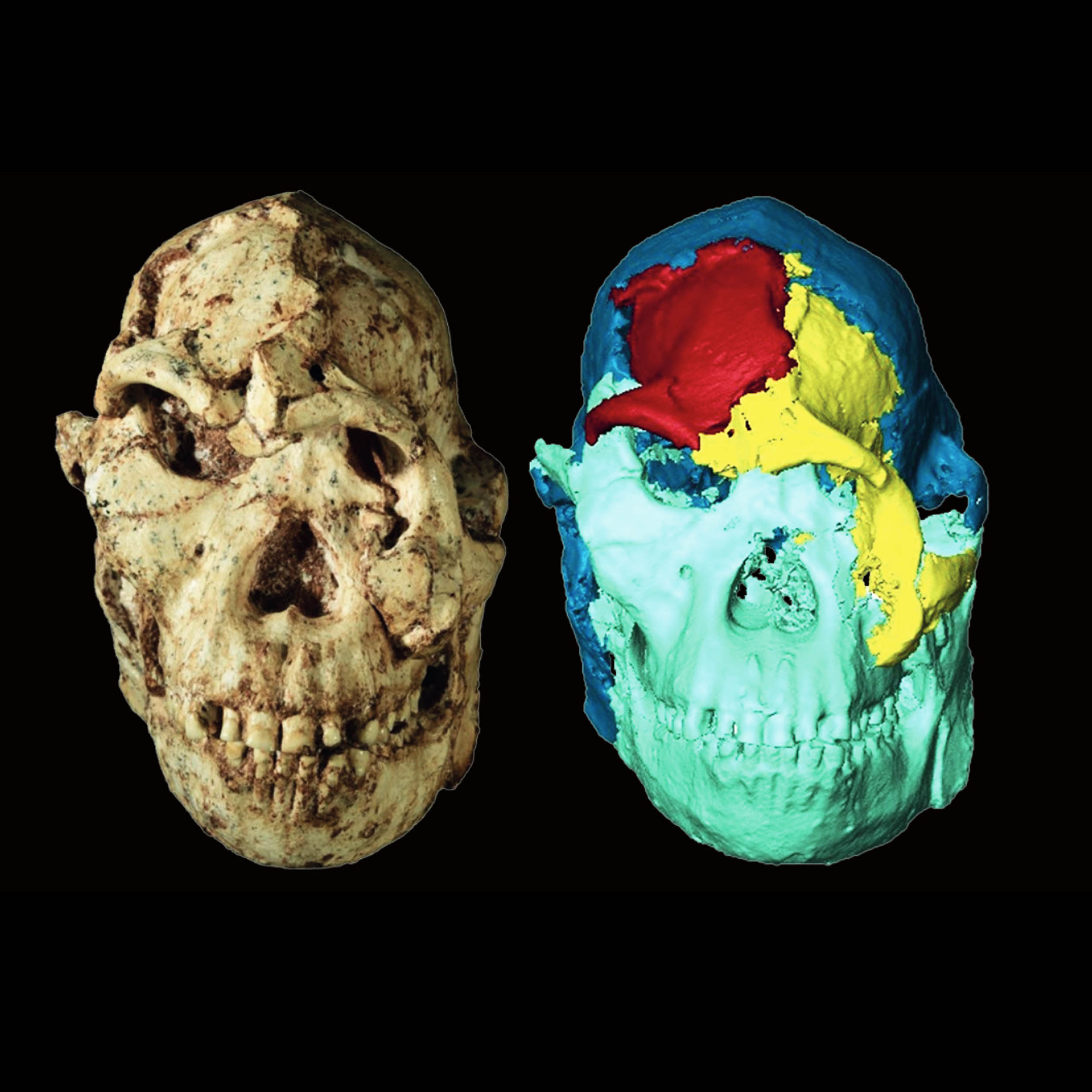
A 3.7-million-year-old face reveals surprising links across Africa
A new digital reconstruction of an ancient fossil’s face is reshaping how scientists think about early human relatives in Africa.
By virtually “undoing” distortions caused by millions of years underground, researchers have produced one of…
Continue Reading