Rare harbour porpoise mating behaviour has been captured by drones off Shetland’s coast.
The footage, taken between 2019 and 2023, provides one of the most detailed records of…

Rare harbour porpoise mating behaviour has been captured by drones off Shetland’s coast.
The footage, taken between 2019 and 2023, provides one of the most detailed records of…

As the US plans a long-duration mission to the Moon under the Artemis Programme, one of the biggest challenges that the scientists face is what astronauts will eat in space. This challenge has led researchers to explore whether crops can be…
Awan, M. J., Rahim, M. S. M., Salim, N., Rehman, A. & Garcia-Zapirain, B. Automated knee mr images segmentation of anterior cruciate ligament tears. Sensors 22, 1552 (2022).
Putera, K. H. et…

Historically, dinosaur ages have been estimated using the growth rings in their bones — one ring per year. But new…

Craig Munns has a large model of a T rex on his desk. He got it with a magazine subscription two decades ago. One day, a few years ago, he was sitting in his study, which was dense with books and yellow sticky notes and posters charting evolution…

Finding Earth-like exoplanets with the composition and ingredients for life as we know it is the Holy Grail of exoplanet hunting. Since the first exoplanets were identified in the 1990s, scientists have pushed the boundaries of finding…
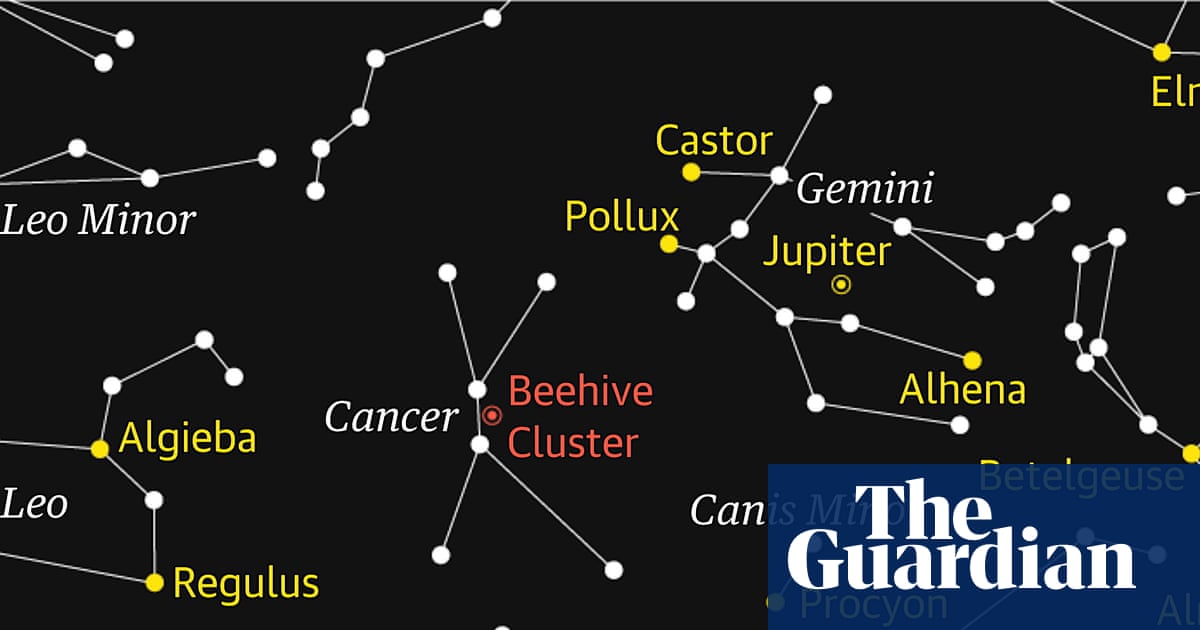
The constellation of Cancer, the crab, is now high in the southern sky during the late evening. While not the easiest constellation to locate because it does not contain any truly bright stars, it does offer a reward for patient observation: the…
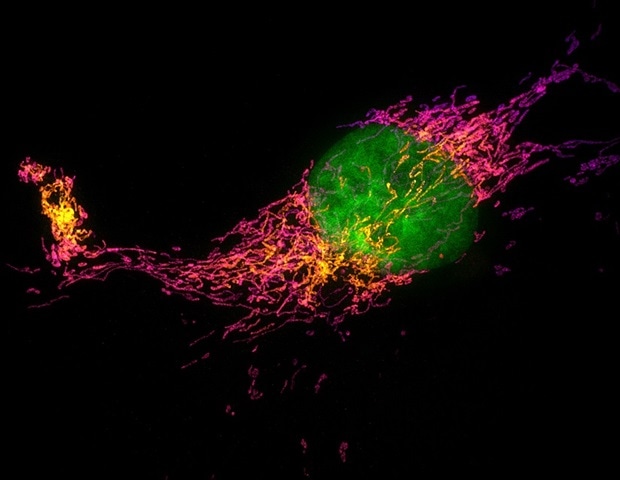
Every time we feel a gentle tap on the skin, specialized nerve cells convert that physical force into an electrical signal that the brain can interpret as touch. While scientists have long known that a protein called PIEZO2 acts as…

by Clarence Oxford
Los Angeles CA (SPX) Mar 09, 2026
NASA says it has finally found the “back to basics” recipe to get Americans back on the Moon by 2028. A new intermediate mission, standardized hardware, a faster launch cadence: on…
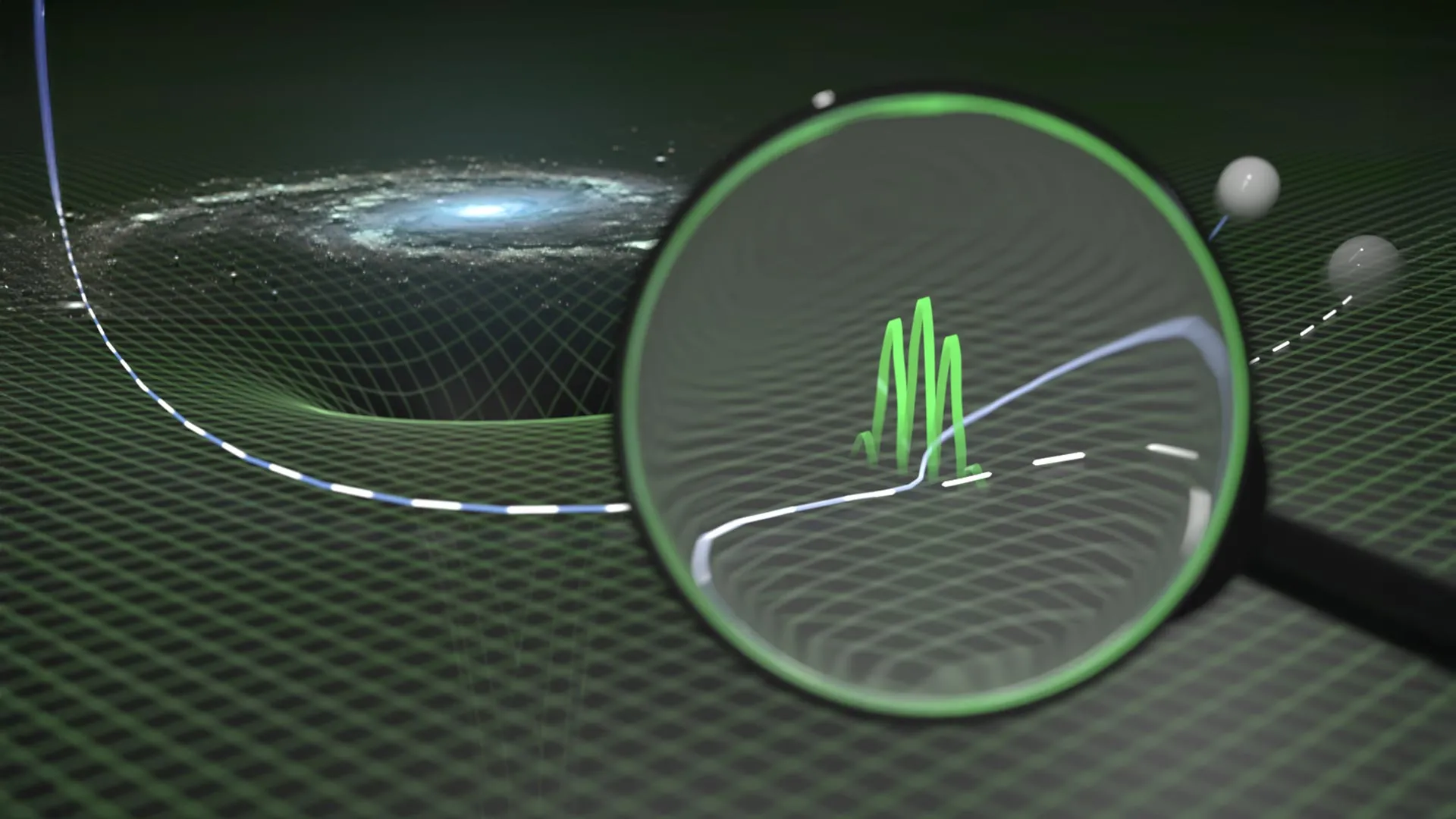
One of the biggest unsolved challenges in modern physics is bringing together two powerful theories that describe very different parts of reality. Quantum theory explains the behavior of extremely small particles with remarkable precision….