Qaasid News
Download Our App
Latest News from Pakistan
PAF jets give spectacular aerial salute to Kazakhstan President – RADIO PAKISTAN
February 3, 2026
Turkey determined to take relations with Saudi Arabia to higher level, Erdogan tells crown prince – Reuters
February 3, 2026
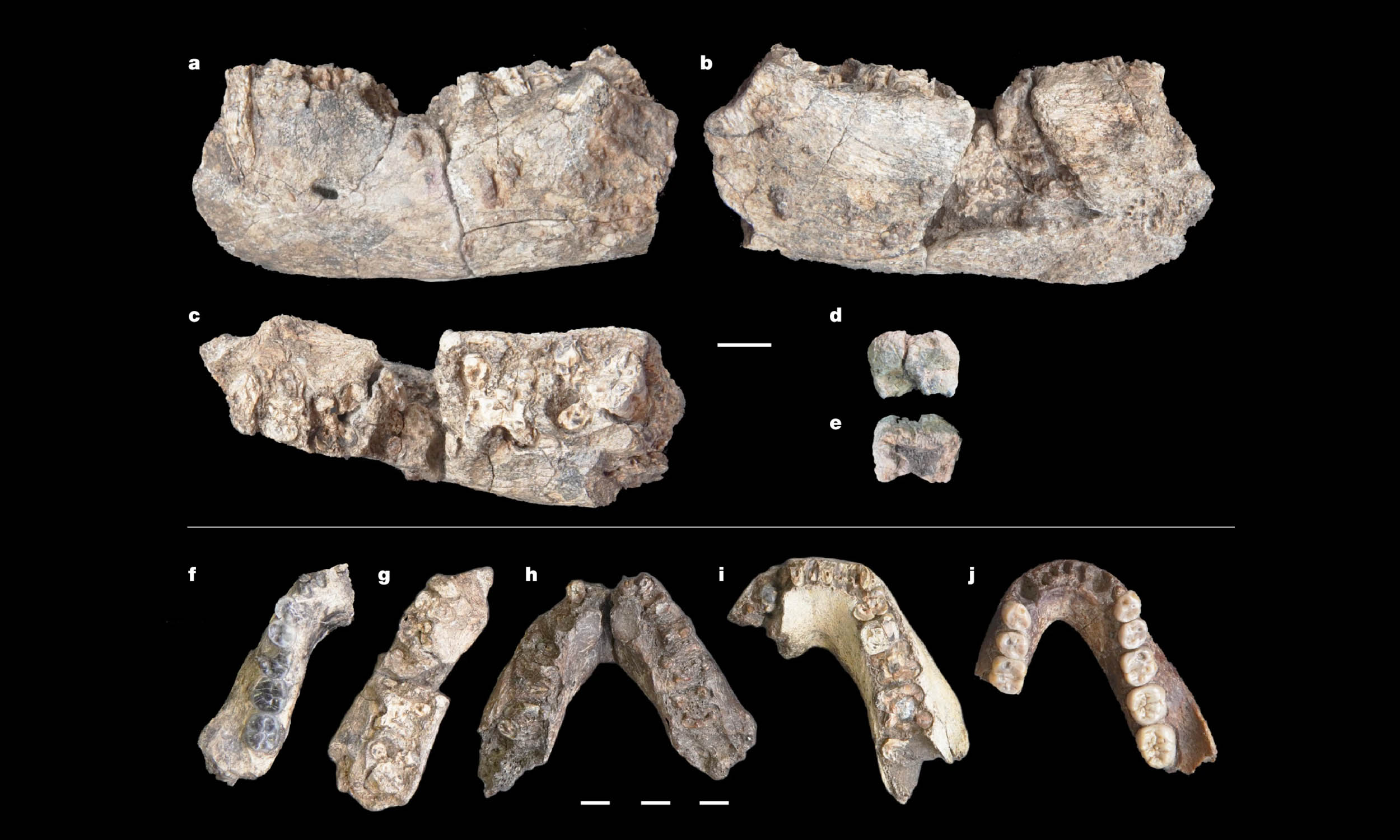
2.6-million-year-old Paranthropus jawbone changes human timeline
February 3, 2026
The Minecraft Safety Council | Minecraft
February 3, 2026
Paul Elliott obituary | Theatre
February 3, 2026
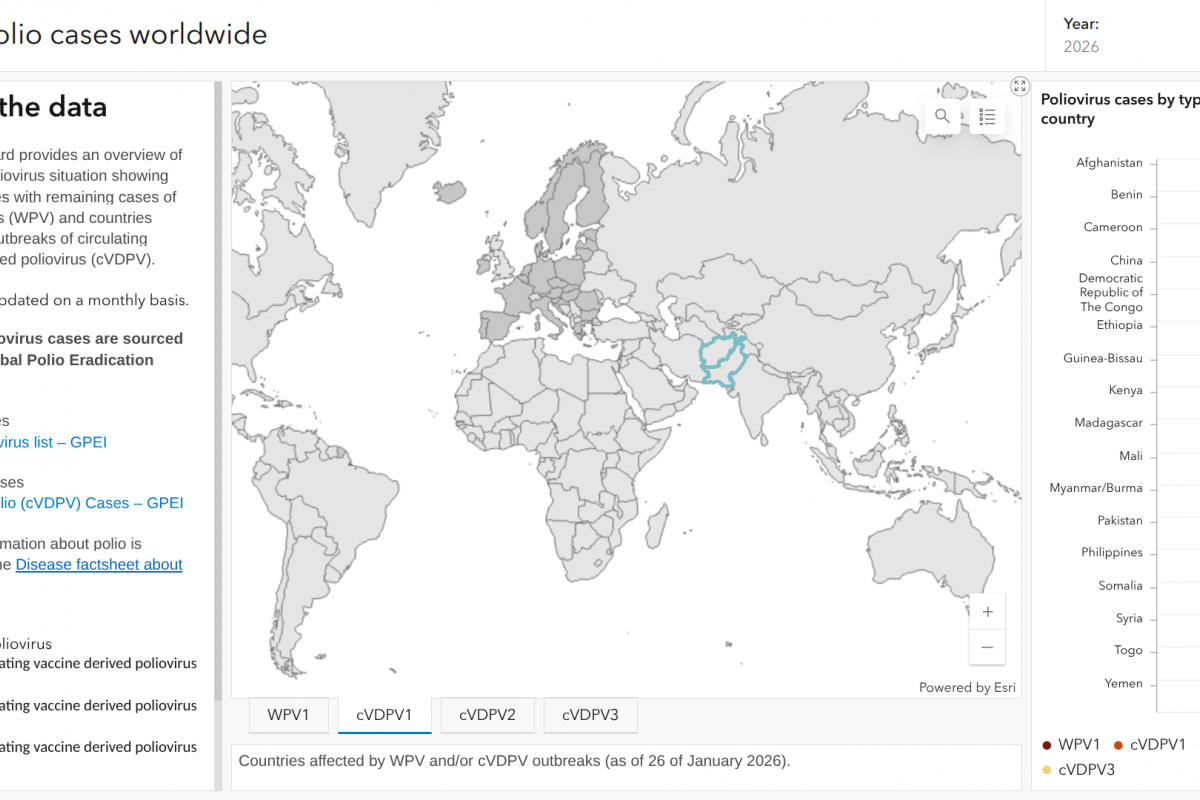
Polio Detections in 2026 Highlight Persistent Global Health Threat — Vax-Before-Travel
February 3, 2026
How giant ‘Blobs’ of rock have influenced Earth’s magnetic field for millions of years – new research
February 3, 2026
Astra-Langchain4j Achieves LLM Integration With Three Agentic AI Implementations
February 3, 2026
Lord of the Dance: Show 'will go on' as Flatley obtains emergency injunction – BBC
February 3, 2026
Exclusive: US shoots down Iranian drone approaching aircraft carrier, official says – Reuters
February 3, 2026