Qaasid News
Download Our App
Latest News from Pakistan
Trump tells Iran to make nuclear deal or next attack will be 'far worse' – Reuters
January 28, 2026
M24 – Motorsport museum set to open on 28 May 2026
January 28, 2026
New nanomaterial kills cancer cells via oxidative stress
January 28, 2026
Starting 5: Sixers roll, Clippers stay hot & a look ahead to Wednesday’s slate
January 28, 2026
Maryam Nawaz’s Governance Model: Delivery, Visibility, and Reform in Punjab
January 28, 2026
From $11B in 2025 Payouts to What We’re Building for Artists in 2026 — Spotify
January 28, 2026
Global Blog | Motorola introduces the moto g77 and moto g67
January 28, 2026
Trump warns Iran 'time is running out' as US military builds up in Gulf – BBC
January 28, 2026
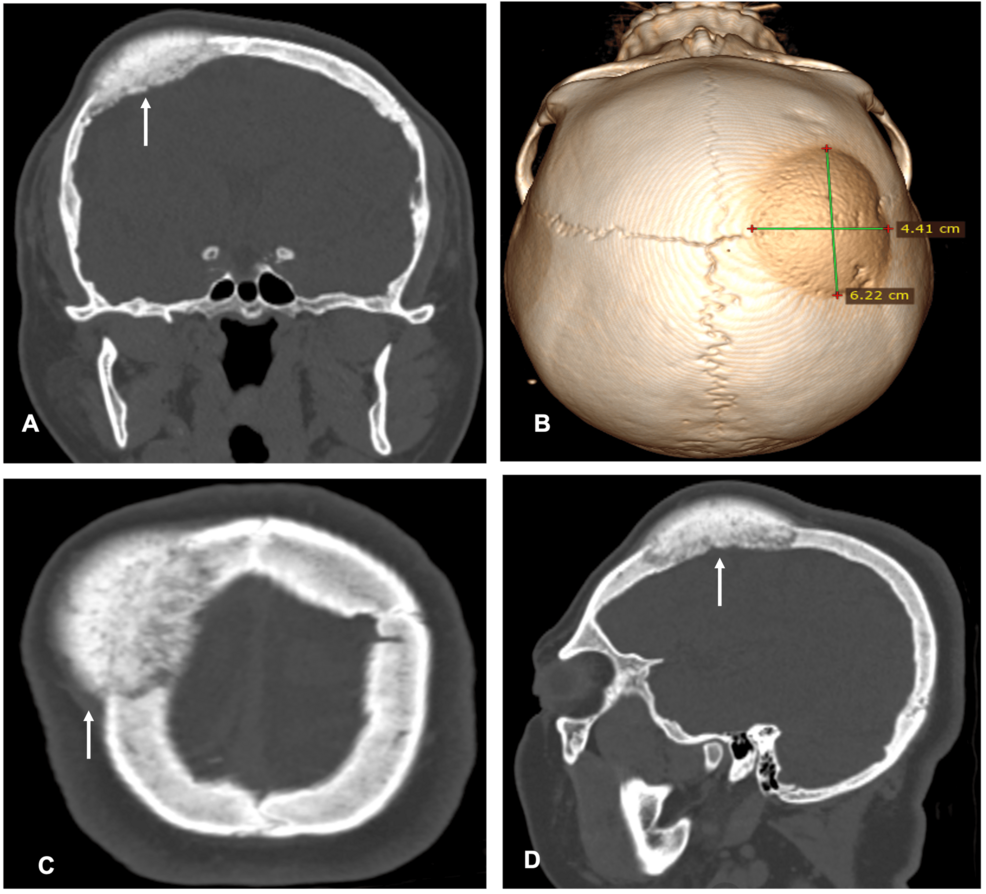
Single-Stage Resection and Reconstruction of Intraosseous Meningioma: The First Reported Case at Our Hospital
January 28, 2026
Immigration espresso shot – New Year Special – Dentons
January 28, 2026