Qaasid News
Download Our App
Latest News from Pakistan
Your browser is not supported
January 12, 2026
Pre-order Lego Pokémon sets today featuring Pikachu, Eevee, Charizard
January 12, 2026
Should You Buy a Folding Phone in 2026?
January 12, 2026
What the surge in gold and silver to fresh records says about the mindset of investors to start 2026
January 12, 2026
Study: Late Ordovician Mass Extinction Cleared Way for First Fishes
January 12, 2026
CSS at Scale With StyleX
January 12, 2026
Recognizing Blood Clots in Unexpected Places
January 12, 2026
Tiny RNA molecules in sperm, big impact on baby health
January 12, 2026
CSULB art grads illustrate the storyboards that bring Hollywood blockbusters to life
January 12, 2026
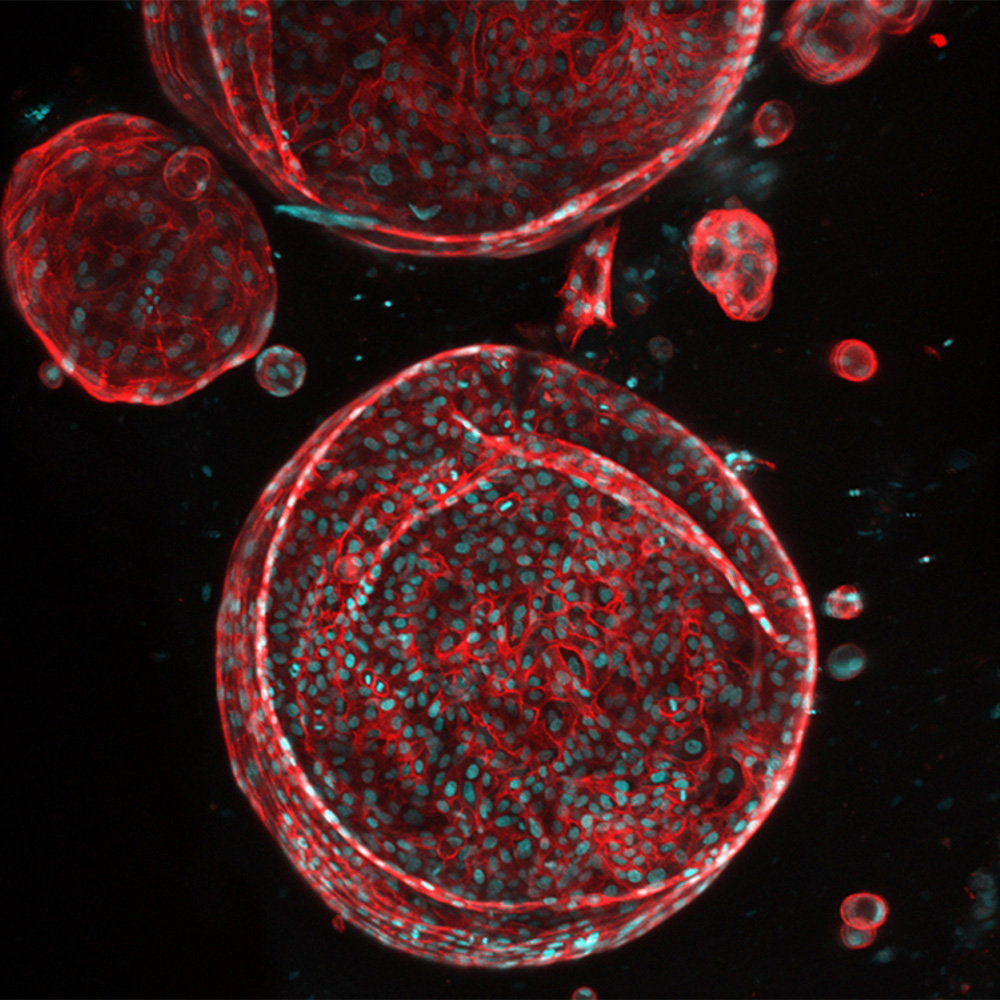
UTSW receives ARPA-H award to create functioning artificial liver: Newsroom
January 12, 2026