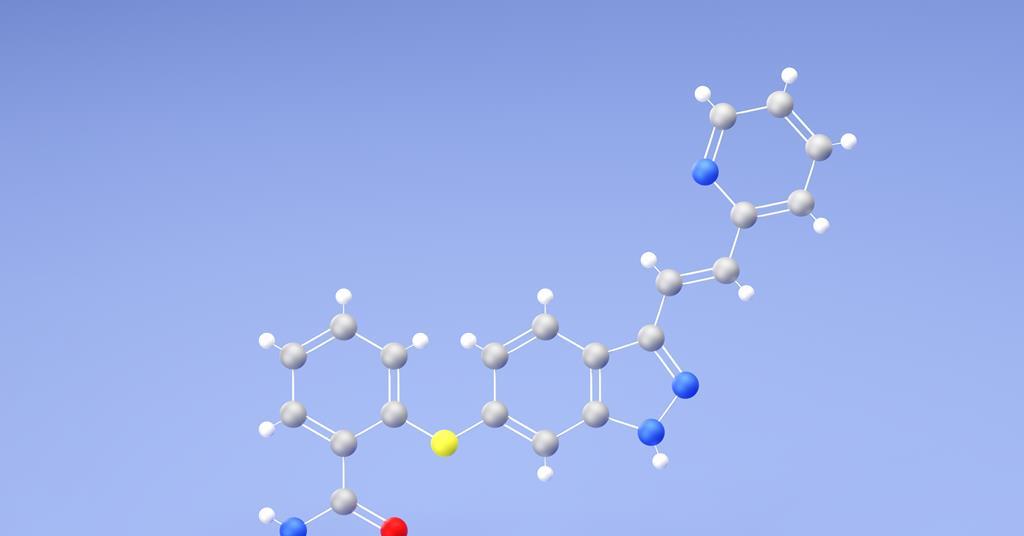
‘Crystal structure prediction is, after years of hard work by many, many groups, finally reaching the point where it’s going to have large impacts in organic materials.’ So says Gregory Beran, at the University of California Riverside in…
Crystal structure prediction method successfully models axitinib polymorphs and salt formation