When microscopic particles of sand, ash, or dust collide in the air, they often exchange a tiny electrical…
Category: 7. Science
-

Scientists Finally Solved a Massive Mystery About Static Electricity Using Acoustic Levitation
Levitating matter with sound. Experimental setup with an acoustically levitated particle of silica. Credit: Thomas Zauner/ISTA -

How Open NASA Data on Comet 3I/ATLAS Will Power Tomorrow’s Discoveries
The interstellar comet 3I/ATLAS will soon leave our solar system, never to return, but the observations of the comet will live on in NASA’s public data archives. More than a dozen NASA science missions turned their instruments to observe…
Continue Reading
-
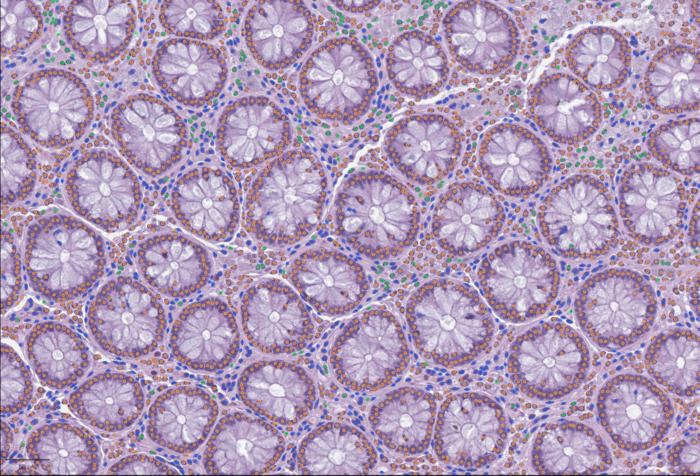
Open Framework for Integrating Whole-Slide and Molecular Data
Cell detection with LazySlide in human colon tissue: Immune cells (green), connective tissue cells (blue) and epithelial cells (orange) [Yimin Zheng] Histopathological data remain one of the most trusted tools in science when…
Continue Reading
-
Molecular Adaptations And Engineering Of Extremophiles For Synthetic Biology And Biotechnological Applications – astrobiology.com
- Molecular Adaptations And Engineering Of Extremophiles For Synthetic Biology And Biotechnological Applications astrobiology.com
- Impact Survival of Microbes Highlights the Feasibility of Panspermia Avi Loeb – Medium
- Scientists Think the Most…
Continue Reading
-

SpaceX Now Has 10,000 Starlink Satellites In Space — And Polymarket Is Betting The IPO Is Coming Soon
Elon Musk’s SpaceX put more than 10,000 active Starlink satellites into orbit this week with back-to-back Falcon 9 launches.
The milestone matters because Starlink is the revenue engine behind what may be the largest IPO in history, and…
Continue Reading
-

Zebra finches reply faster when they hear a familiar bird call
Researchers have found that zebra finch brains show stronger and longer-lasting activity in neurons that control vocal timing when the caller is familiar compared to when it is unfamiliar.
The result links social recognition directly to the neural…
Continue Reading
-

Scientists Bring Mouse Brains Back to Life After “Cryosleep” Deep Freeze
The concept of “cryosleep” — spending prolonged periods of time in suspended animation in…
Continue Reading
-
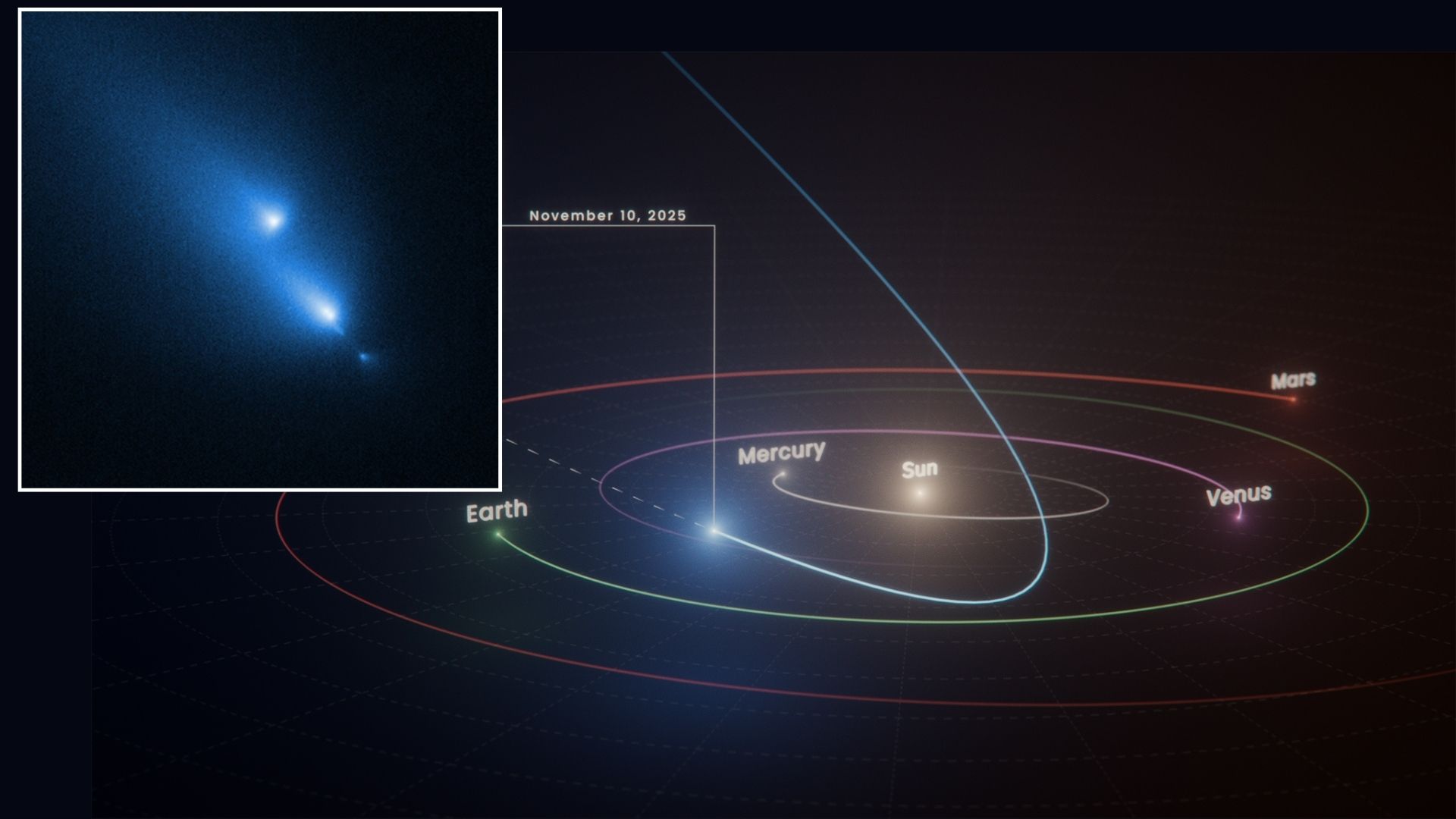
Hubble Space Telescope accidentally witnesses comet C/2025 K1 (ATLAS) breaking apart
NASA’s Hubble Space Telescope has captured a rare cosmic moment: a comet breaking apart in real time.
During its routine imaging of the universe, the space telescope spotted an unexpected object called C/2025 K1 (ATLAS), or comet K1 for short….
Continue Reading
-

Perseverance’s radar revealed ancient subsurface river delta on Mars
When NASA’s Perseverance rover landed in Jezero Crater in 2021, its primary mission was to scour the remnants of a dried-up Martian lakebed for signs of ancient life. Scientists have…
Continue Reading
